|
SUBFAMILY
CHARACTERISTICS
Adults
The prefix sticto- in the subfamily name means dappled, mottled
or spotted. The majority of species have most finely detailed markings to the
forewings, often a clearly etched reticulate patterning. Many genera contain
species with a distinctive hyaline basal half to the hindwing, particularly Stictoptera
Guenée, Aegilia Walker and Lophoptera Guenée.
The wing venation is as illustrated in Figs. 8-12, and is constant
within the subfamily except for secondary sexual modifications in the male of Sigmuncus
arcuata Hampson (Fig. 11).
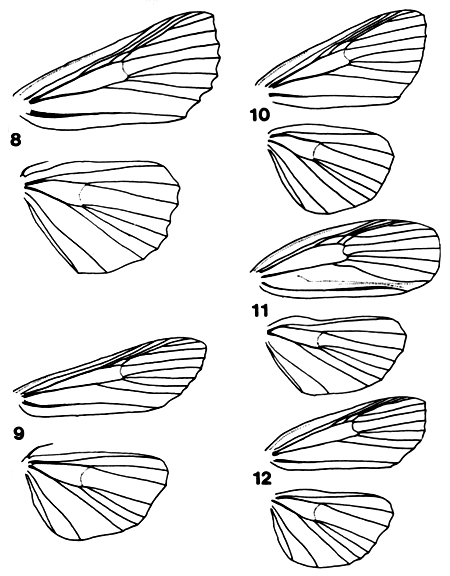
Figures 8-12. Venation of Stictopterinae; 8 ( ) and 9
( ) and 9
( ) illustrate the difference in frenulum curvature mentioned in the text. 8. Stictoptera
signifera, 9. Gyrtona lapidarioides, 10. Diascoides ferruginea, 11 () and 12 () Sigmuncus
arcuata. ) illustrate the difference in frenulum curvature mentioned in the text. 8. Stictoptera
signifera, 9. Gyrtona lapidarioides, 10. Diascoides ferruginea, 11 () and 12 () Sigmuncus
arcuata.
The antennae of both sexes are filiform, sometimes finely ciliate.
Ciliation appears to be more common in, and is perhaps exclusive to, the quartet
of genera that includes Gyrtona Walker in Fig. 13.
The frenulum in the female is reduced to a single spine; it tends to
show its greatest curvature over the basal half whereas that of the male is more
evenly curved, often with a slight angle centrally.
The main flap of the counter-tympanal hood is usually longer than its
base and, in dried specimens, directed anteriorly over the tympanum; it is
overlapped by the concavity of a fan of narrow, lenticular scales arising from
dorsally to the tympanum on the metathorax. The small flap ventral to the
counter-tympanal hood (Figs. 155, 157) forms a shallow concavity into which the
ventral part of the base of the hood extends. A tuft of fine scales extends
obliquely in a posterior ventral direction from anterior to the tympanum as in
the Euteliinae but not to such a pronounced extent.
The basal sternite of the abdomen has strengthening infolds (grooves
from the exterior) extending posteriorly from the apophyses (Figs. 155, 157)
rather than interior flanges as in the Euteliinae. Such grooves are seen in many
other noctuid subfamilies. The male eighth sternite typically bears a pair of
coremata.
In the male genitalia there is a small sclerotised lobe just inside the
ductus ejaculatorius from the aedeagus vesica, a character shared with the
Euteliinae but as yet not sought rigorously in other subfamilies. The valves are
usually rather narrow (or bifid with both arms narrow), often with a process
from the base of the costa (e.g. in many Lophoptera); the harpe, when
present, is usually small, subbasal on the sacculus and ampullate (e.g. in Savoca
Walker and Aegilia indescribens Prout). In one lineage the valve is
divided into almost equal costal and saccular arms, sometimes united by a
flexible folded membrane (Gyrtona and allies).
The bursa copulatrix of the female has a single scobinate signum (absent
in a few cases), the scobination appearing in most instances to be a
concentration and increased sclerotisation of a more general scobination of the
bursa. The ovipositor lobes have numerous setae on elevated bases, and the
margins are usually somewhat divided to give a ‘frayed’ appearance; the
lobes are mostly semicircular, forming a disc together, but being acute in only
a few species such as Lophoptera pallibasis Holloway and allies, and Gyrtona
lapidarioides sp. n.
Species tend to rest with wings folded at an acute angle or, in Stictoptera
and the more narrow-winged species, with them rolled round the body giving
the appearance of a short, broken twig.
Larvae
Gardner (1948a) described the larva as having a complete set of four
equal pairs of prolegs with uniordinal crochets, variably homoideous (Lophoptera)
and heteroideous (Odontodes). The mandible has no internal tooth and
the head has the frons higher than wide, acute at the apex, not shorter than the
coronal suture. The body skin is smooth.
These observations were based on Lophoptera and Odontodes, for
which Gardner also provided generic descriptions. There was no reference to any
member of the Gyrtona/Stictoptera lineage. Also the sample of three
species from two genera is probably too small for valid generalisations.
Pupae
The pupa is of typical noctuid shape but the cremaster is of constant
type within the subfamily (observed in Stictoptera, Savoca, Aegilia and Lophoptera
species reared by Bell, and for Lophoptera and Odontodes by
Gardner (1948b)) and may prove diagnostic, being somewhat unusual within the
Noctuidae. It was illustrated by Gardner.
The cremaster consists of a Y-shaped arrangement of two conical spines,
fused over a basal stalk twice as wide and one third as long as the arms of the
Y. The arms splay out at an obtuse angle, slightly outcurved. The Y is flanked
on each side by two or three small hooklets.
Pupation is invariably in the soil surface in an ovoid silken cocoon
incorporating particles of soil.
Relationships between genera
In Fig. 13 a tentative scheme of relationships between the eight genera
is presented, based on the following presumed apomorphic character-states:
|
1. |
Pupal cremaster a distinctive Y-shape.
Female
frenulum reduced to a single spine.
‘Frayed’,
coarsely setose ovipositor lobes arranged in a disc.
Counter-tympanal
hood structure as described above.
General
wing facies characteristics as described.
|
| 2.
|
Male
valve divided into two more or less equal slender arms (cf. valves entire
with only short costal processes).
Valve costal processes tend to be absent (except in some species of Savoca
and Aegilia describens).
|
|
3. |
The male saccus is more or less trilobed.
A
membrane links the valve arms (though this may be a plesiomorphy relative to
complete loss in Gyrtona).
Loss
of saccular harpe (except in Aegilia indescribens where there is a basal
structure that may be homologous with the harpe in Gyrtona).
|
|
4. |
Female 8th sternite much more heavily developed than the tergite, with apophyses
usually short and broad, though not so markedly so in Diascoides and Sigmuncus
(cf. the 8th segment almost a complete ring with normal, long, slender
apophyses).
Ciliation
of male antennae very much reduced over basal half (assuming general, obvious
ciliation is plesiomorphic). |
|
5. |
Female signum with scobination distally directed.
Appendix
bursae well developed, often almost equal in size to bursa.
Apophyses
of female 8th segment short and broad.
Apomorphic characters for each genus are given in the generic
descriptions.
|
HOST-PLANT RELATIONSHIPS AND ECONOMIC IMPORTANCE
The host-plant relationships within the subfamily are very interesting as
far as can be determined from the sparse data available.
All records for the Stictoptera/Gyrtona lineage, for the genera Stictoptera,
Aegilia and Savoca, are from genera of the family Guttiferae such as Mesua,
Garcinia and Calophyllum. Records from this family have been made in
India (Mathur 1942; Gardner 1948a; Browne 1968; Bell, MS), S.E. Asia and
Sundaland (Pholboon 1965; CIE records; Dr Tho Yow Pong, pers. comm.), Fiji
(Robinson 1975) and Hawaii (Beardsley 1982). No host-plant for any member of the
genus Gyrtona could be located but the abundance of members of this genus
in montane samples during the Mulu survey and the frequency in heath forest
collections leads to the tentative prediction that the hosts may also be in the
Guttiferae: this family provides an important component of Bornean lower and
upper montane forest and is also strongly represented in heath (kerangas) forests. Species of Stictoptera,
Aegilia and Savoca were also found to be somewhat commoner in montane
localities but not in the kerangas samples.
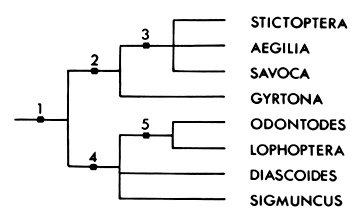
Figure 13. Diagram of relationships of stictopterine genera. The numbers refer to
apomorphic states of characters used to define the subgroups as listed in the
text.
Records from the Lophoptera lineage are few but show a high
frequency in the family Dipterocarpaceae: Odontodes aleuca Guenée
from Shorea (Gardner 1948a); Lophoptera parallelnotata sp.
n. from
Shorea (P. Becker, pers. comm.); Lophoptera illucida Walker from Shorea
(Gardner 1948a) and Hopea (Bell, MS). Two species have a recorded
host range that extends to other families: Lophoptera squammigera Guenée
is known from Shorea but also Grewia (Tiliaceae) and Mallotus and
Briedelia (Euphorbiaceae) (Mathur 1942; Gardner 1948a; Bell, MS); a
specimen tentatively identified as Lophoptera hemithyris Hampson was
reared from Glochidion (Euphorbiaceae) in Fiji (Robinson 1975).
A preliminary survey of host-plant records for Indo-Australian
macrolepidoptera has indicated a very low number of species feeding on
Dipterocarpaceae relative to the importance of the family in south-east Asian
lowland rainforests. Most records for the family tend to be of polyphagous
lepidopteran species from families such as Lymantriidae, Arctiidae and
Lasiocampidae (Mathur et al. 1954-1960) which themselves contain a high
proportion of polyphagous taxa. Hence the incidence of specific records in the Lophoptera
lineage is of great interest and merits further investigation. Many species
of this lineage are more strongly represented in the lowland forest samples of
the Mulu survey than are members of the Stictoptera lineage, though there
are also some species that appear to be most abundant in montane samples,
suggesting that other plant families are involved as hosts.
The geography of the two lineages also reflects the geography and
diversity of the two plant families through the Indo-Australian tropics to some
extent, as discussed in the next section.
The genera of Guttiferae mentioned include many trees of economic
importance. Several produce valuable timber, species of Calophyllum yield
useful resins and oils, the flowers of some Mesua yield a perfume and the
genus Garcinia can have edible fruits such as the mangosteen, G. mangostana.
Species of the Stictoptera lineage, especially of Stictoptera itself,
may achieve population levels sufficient to attain pest status, defoliating
guttiferous trees.
The effect of members of the Lophoptera lineage as defoliators of
Dipterocarpaceae has yet to be gauged, but it is conceivable that both they and
members of the Stictoptera lineage could affect the composition of forest
regrowth by grazing tree seedlings.
ZOOGEOGRAPHY
Species and species complexes of the Stictoptera lineage tend on
the whole to be wider ranging in the Indo-Australian tropics than are those of
the Lophoptera lineage. The Stictoptera lineage has, in Borneo, 13
species out 34 extending from the Oriental tropics to the Moluccas or New Guinea
whereas there are 12 out of 55 in the Lophoptera lineage, though this
latter total increases slightly if close-knit species groups are included. There
are several Gyrtona endemic to the south-west Pacific (Robinson 1975;
under both Gyrtona and Nigramma); Savoca divitalis Walker ranges
into Polynesia; a species complex in Aegilia reaches Samoa and has
endemics in several south-west Pacific archipelagos; there are endemic and local
species of Stictoptera in the south-west Pacific (Robinson 1975; Holloway
1979), all of which, from a consideration of the range of forms in each, are
allied to cucullioides Guenee and signifera Walker.
In the Lophoptera lineage Odontodes extends to the
Bismarcks and Sigmuncus to New Guinea. Diascoides is purely
Oriental. Lophoptera is extremely diverse in Sundaland, moderately so in
mainland tropical Asia, and has several species in New Guinea. Further east
there are very few species: in New Caledonia there is only vittigera Hampson
(misidentified by Holloway (1979) as squammigera; see the account of squammigera
below) and hemithyris, and in Fiji only the latter. Both these
species probably feed on Euphorbiaceae (vittigera is closely allied to squammigera).
It remains to be seen whether the correlation of species-richness in Lophoptera
with that of the Dipterocarpaceae is more than coincidental.
Endemism in Borneo is moderate with six species in the Stictoptera lineage,
five of which are commoner or only taken in upper montane localities, and eight
in the Lophoptera lineage, three of which have only been taken in upper
montane localities, the rest being lowland taxa.
SYSTEMATIC ACCOUNT
All relevant generic type species were examined and both sexes of each
dissected as for the Euteliinae. Again, the result has been a virtual revision
of the Indo-Australian members of the subfamily and many changes, such as the
subordination of Nigramma Walker to Gyrtona and the transfer of
many species from Lophoptera to Savoca and Gyrtona, and
from Gyrtona to Savoca and Lophoptera. Two new genera, Diascoides
and Sigmuncus, are described.
>>Forward
<<Return to Contents page
|