|
This
tribe, as circumscribed for this work (a broader concept is adopted by Fibiger
(2003), who treated the grouping here as a subtribe), was investigated in detail
by Mitter & Silverfine (1988); it includes in synonymy the Audeinae of
Wiltshire (1990), a nomen nudum according to Speidel & Naumann
(2005).
The
definitive features in the female genitalia are: elongated, narrow, tapering
ovipositor lobes with a longitudinal band of sclerotisation extending through
them from the apophyses (but see also the Catephiini on p. 84, Ericeia
Walker
on p. 116, Dinumma Walker on p. 242, Hamodes Guen�e
on p. 267 and Chilkasa Swinhoe on p. 291); an ostium set well anterior at
the end of a deep cleft with the seventh sternite. The posterior margin of the
sternite is represented by lobes at the end of the cleft, but it is not clear
that these are homologous with the bilobed condition of the antevaginal plate
seen in other tribes such as the Ophiusini, given that it is entire in some
genera, covering the ostium. The cleft seventh sternite is seen also in the
putative sister-genus Metatacha Hampson.
The male
genitalia were found by Mitter & Silverfine to be difficult to interpret.
The features are generally as in the core Catocalinae as discussed on p. 17, but
the valves distally are divided into a strongly sclerotised costa and a
membranous to corematous distal part suspended from the costa. There is usually
a single, finger-like process from the sclerotised sacculus; its relationship to
features in a similar position in, say, the Ophiusini (p. 40) is unclear. The
valves often show bilateral asymmetry. The juxta is of the inverted �Y�
type, and the structures of the uncus, scaphium and aedeagus are consistent with
the general condition of the core catocalines.
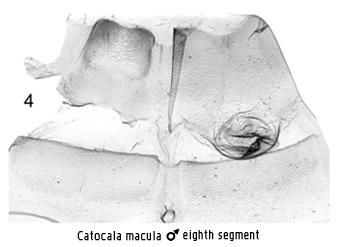
However,
unlike most core catocalines, the male eighth segment shows vestiges of the
framed corematous condition in many taxa (e.g. Fig 4) both in the sternite and
the tergite, and in some taxa its development is more obvious (Figs 25 and 26 in
Mitter & Silverfine (1988)). If the interpretation of lack of modification
of this segment as a synapomorphy for a core catocaline group is valid, then the
group of genera including Catocala Schrank
might be placed relatively basally within this core group, perhaps also
supported by the somewhat different structure of the ostium and seventh sternite
in the female. Given these uncertainties, it would be preferable to retain this
narrow concept of the Catocalini separate from other core catocaline tribes for
the time being.
Larval
and pupal features (e.g. a bloom) support a general association of the group
with other core catocalines as discussed on p. 22 and p. 24, though reduction of
the prolegs is only slight.
Mitter
& Silverfine (1988) presented a phylogeny for the group that separated Catocala
and
Ulotrichopus
Wallengren
from a predominantly African clade that included Audea Walker,
Crypsotidia
Rothschild
and Hypotacha
Hampson.
Within the Catocala clade, Ulotrichopus species
showed monophyly but were nested within Catocala.
Recognition of Ulotrichopus
as
a good genus (e.g. as in Kobes (1985), Poole (1989) and Fibiger (2003)) would
render Catocala paraphyletic in this system, therefore Ulotrichopus
is
included as a synonym of Catocala, an arrangement also adopted by Fu & Tzoo
(2002).
>>Forward <<Return to Content Page
|
