|
The Lasiocampidae
are reasonably diverse in the Old World tropics, less so in the new. They have
moderate representation in the Holartic and Australasia, but are not
represented east of Papua New Guinea. They are generally large species with deep
forewings, and hindwings at least two thirds of the area of the forewing, often
rounded; however there are several genera in Africa and the Indo-Australian
tropics where the forewings are narrow, apically produced, and the hindwings
relatively much smaller, the abdomen protruding well beyond them. These have
been suggested in the introductory section to fill the niche occupied in the
Neotropics by similarly Saturniidae. Sexual dimorphism is usually marked.
Adults
The tongue is
vestigial or absent. The male antennae are strongly bipectinate, though this is
sometimes reduced over the distal portion; those of the female are usually less
so. The frenulum is absent, the humeral lobe of the hindwing being expanded and
often containing humeral veins. Patterning on the hindwing is costal (dorsal or
complete in other bombycoids).
In the forewing
venation R2 and R3 have an common stalk, as do R.5 and M1, usually
with R4 branching off from this stalk more basally and anteriorly. A few
Oriental taxa have the latter triplet with R4 and R5 bifurcating distally,
with M1 arising more basally and posteriorly as in the Mimallonidae (see Fig.
2). The anterior cubital vein in both wings is strong with its two branches and
M2 and M3 arising from it in a quadrifid array. On the hindwing Sc and Rs
anastomose for a short distance subbasally, forming a basal or humeral cell
anterior to the main cell (Fig. 3). This basal cell becomes considerably
expanded in the Gastropachinae (Fig.3).
In the male
genitalia the saccus is often associated with a complex plate or two elongate
arms that usurp the function of the rather reduced valves; this structure
was termed the cubile by Lajonquiere (1968). The aedeagus is often short,
sometimes with a ventral spur, and the vesica is frequently scobinate.
The uncus is reduced, the gnathus lost, and the
tegumen often weak except in one subfamily restricted to the New World (see
below).
Larvae
The larvae
are densely invested with secondary setae and appear very hairy. They are
cylindrical or rather flattened, usually with lateral lappets or protruberances
below the line of the spiracles on each segment, the prothorax having a second
protruberance above the first. In many genera there are also transverse saddles
of densely packed short setae dorsally on T2 and T3 (see Plate 19; Gastropacha
pardale Walker). If one attempts to pick a larva up, the dorsal part of the
thorax is brought strongly back against the hand (or attacker), leaving the
short setae of the thoracic saddles embedded in it (Tho Yow Pong, pers.
comm.). The cocoon is well developed.
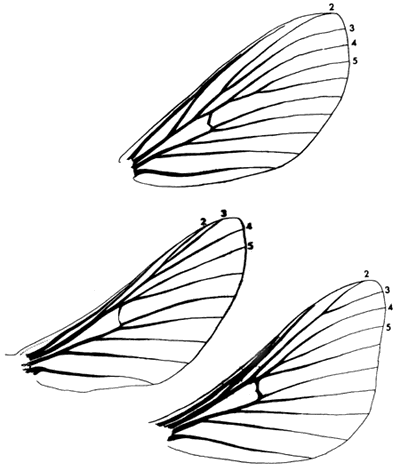
Figure 2.
Forewing venation of Lasiocampidae: top, typical configuration (Paralebeda);
centre genus Bhima, bottom, 'mimallonoid' venation shown by Alompra. Radial veins 2 to 5
are indicated by numbers.
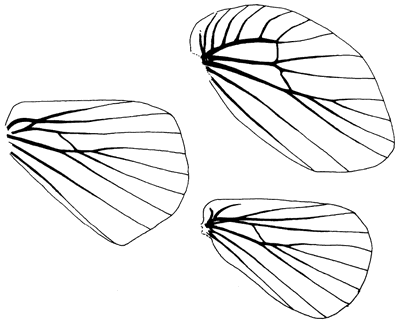
Figure 3.
Hindwing venation of Lasiocampidae: left, most frequent South-east Asian
configuration (paralebeda); top right, gastropachine configuration (Gastropacha
leopoldi), with numerous humeral veins anterior to Sc, and a major additional
cell between Sc and Rs; bottom right, Syrastrena, with rather basal
humeral veins, otherwise similar to Paralebeda.
Subfamilies, and
groupings of Oriental genera
Franclemont (1973)
reviewed the subfamilial classification, recognising three subfamilies in the
New World. The Macromphaliinae may well be restricted to that region,
and appear somewhat plesiomorphic in male genitalia characters: the uncus,
gnathus or socii are usually present, and there is no cubile. There are three
small Africa subfamilies, Chondrosteginae, Chionopsychinae and Archaeopachinae,
and a fourth, Gonometinae, that Franclemont suggested might be included in the
Gastropachinae. The
Gastropachinae and Lasiocampinae are the other two New World subfamilies and
occur worldwide. Only these two are represented in the Oriental fauna, and their
validity needs investigation.
The
Gastropachinae are defined by massive expansion of the humeral cell and the
presence of a sequence of humeral veins arising from Sc on its anterior
margin (Fig. 3). Yet the presence of a cubile and the double structure of
the ante- and postmedial fasciae of the forewing suggest they may represent just
a section of a grouping within the Lasiocampinae.
The genera discussed first in the systematic section all have long, rather
slender cubile arms; amongst them are the taxa with the most extreme development
of the narrow forewing and reduced hindwing. Paraleda and Streblote appear to have
single fasciae, those of the rest and the Gastropachinae being double. The
Gastropachinae, Bhima, Metanastria and Lebeda have the discal spot
between the doubled fasciae; in Suana and Kunugia it is more basal, between the
antemedials, and it Lajonquierea, with a very short cell, it is even more basal
still. Apart from the Gastropachinae none of these genera has humeral veins.
Thoracic larval saddles tend to be well developed in these genera, but not in
the few instances where larvae of the next group are known.
Single fasciae
occur in all the genera from Arguda to Trabala in the systematic section. The
cubile is broad, not divided into arms in most genera, reduced in some, absent
in Trabala. Some have one or two humeral veins, but these arise from the base of
the humeral cell and are connate when there is more than one (Fig. 3). The small
complex of genera with mimallonoid venation (p. 11) has such humeral veins.
Host-plant
preferences
Most
Lasiocampidae are polyphagous, the 12 plant families most frequently noted being listed in Table 4. Leguminosae and Myrtaceae are most often
utilised. 27 other families have been recorded once or twice as lasiocampid
host-plants in the region: Anacardiaceae, Annonaceae, Barringtoniaceae,
Bischofiaceae, Bombacaceae Capparidaceae, Casuarinaceae, Combretaceae,
Elaeocarpaceae, Flacourtiaceae, Geraniaceae, Gramineae, Guttiferae, Lythraceae,
Melastomataceae, Meliaceae, Moraceae, Myrsinaceae, Naucleaceae, Polygonaceae,
Punicaceae, Rhamnaceae, Sapindaceae, Sonneratiaceae, Sterculiaceae,
Tamaricaceae, Urticaceae, and the conifer families Cupressaceae and Pinaceae.
Geography and
habitat preference
Of the 60
species known from Borneo, 18 are endemic, 31 Sundanian, three Sundanian and
also recorded from S. China (Metanastria gemella, Syrastrena sumatrana, Trabala
pallida), four are from the N.E. Himalaya and Sundaland, and the remaining four are
widespread in the Oriental tropics.
The genus
Lajonquierea is purely Sundanian, and Arguda, Radhica, Radhica,
Syrastrena,
Trabala and Micropacha are most diverse in Sundaland. Chonopla extends north to
Burma. Most of the other genera are widespread in the Oriental tropics, Euthrix
being also widespread in the Palaearctic and Gastropacha similarly, though also
represented in Africa. Streblote is predominantly Africa, reaching its most
easterly limits in Borneo and the Philippines. The genera Bhima and Takanea are
unusual in being predominantly subtropical, with one outlying species in Borneo
or Sundaland.
To the east,
Sulawesi has a highly endemic fauna, with single species of Lebeda,
Paralebeda,
Suana, Hallicarnia, Odonestis and Trabala, and two of Kunugia. The only
non-endemic is a Gastropacha species that also occurs on the Philippines and in
mainland Asia but has not been recorded from Borneo.
In tropical Australasia, particularly Papua New Guinea, there are taxa of Oriental
affinity such as Cyclophragma (cf. Kunugia), Pseudophyllodes (cf.
Suana),
Pararguda (cf. Arguda) and two genera in the Gastropacha group. In Australia
there is an endemic group that does not appear to have close affinities with the
Oriental fauna. The family does not occur from the Solomons eastwards.
15 species have
only been taken in montane forests, two, Paradoxopla cardinalis and
Trabala
rotundata, only from G. Kinabalu. 30 have only been taken in the lowlands, and
one of these, Streblote helpsi, appear to be restricted to coastal habitats. The
remaining 15 have been taken over a range of altitudes.
Systematic account
The order of
genera follows the currently accepted higher classification, Lasiocampinae
followed by Gastropachinae. The quadrifasciate Lasiocampinae with a strong cubile
are treated first, followed by the bifasciate genera where the cubile is weak or
absent.
<<Back
>>Forward <<Return to Contents page
|
