|
The
Lithosiinae are the largest subfamily of Arctiidae in the Oriental
tropics. In an earlier part of this series (Holloway, 1988; Part 6), it
was suggested that the arctiid subfamilies Syntominae, Thyretinae,
Ctenuchinae, Euchromiinae all showed reduction or fusion of hindwing veins
and therefore might be related. Subsequent work (e.g. Bendib & Minet,
1998; Kitching & Rawlins, 1999; Jacobson & Weller, in press) has
indicated a relationship between (and synonymy of) the last two, but
within the Arctiinae, and between (and synonymy of) the first two, but in
a sister-relationship (Jacobson & Weller) to the Lithosiinae. The
paired pheromone glands of females noted by Holloway (1988) as a potential
autopomorphy for the Arctiidae have been confirmed as such (Bendib &
Minet, 1988; Kitching & Rawlins, 1999).
The sister-relationship
between the Lithosiinae and Syntominae is supported unambiguously by the absence
of the MSD seta on the larval abdomen. In both subfamilies lower plants,
lichens, algae, mosses and liverworts, feature prominently in records of larval
diet (Rawlins, 1984; Kitching & Rawlins, 1999), and records from higher
plants therefore need to be treated with caution, as the larvae may merely be
browsing on lower plants that are epiphytic on the trunks, twigs and even
foliage of higher plants (see below). However, Bendib & Minet (1998)
consider there is stronger support for grouping the Syntominae with the rest of
the Arctiidae, leaving the Lithosiinae as the basal branch.
The systematics of the
subfamily has been relatively neglected since the time of Hampson (e.g. Hampson,
1900), particularly in the tropical centre of its diversity. Birket-Smith (1965)
attempted to bring some order to the African Lithosiinae. In the Oriental
tropics a few individual genera or restricted faunas have been reviewed (e.g.
van Eecke, 1930; Roepke, 1946a, b; Cern�, 1993). Most recently, Bendib &
Minet (1999) have defined the subfamily and attempted a tribal classification as
discussed in the next sections.
Definition of the
subfamily
The subfamily is most
reliably defined on features of larval morphology (Bendib & Minet, 1999): a
mandible modified by a considerably enlarged mola, a basal area specialised for
maceration of lower plant tissue (Gardner, 1943); on the labrum the M1 setal
bases are well separated ventrad, rather than laterad, from those of the M2
setae.
However, for most genera,
particularly in tropical regions, larvae are unknown, hence features of the
adult are necessary to identify members of the subfamily. Bendib & Minet
(1999) note two adult characters of the abdomen: a tendency for the anterior
regions of each segment of A4 to A6 to be unpigmented, usually becoming bluish
with chlorazol black; the postspiracular tergopleural sclerite of A1 is absent.
Both of these features require dissection to observe, and the second may be a
further synapomorphy for the Lithosiinae and Syntominae.
There are a few more
traditional characters (Kitching & Rawlins, 1999; Bendib & Minet, 1999)
that may enable most Oriental Lithosiinae to be recognised: reduced ocelli or
absence thereof (Macrobrochis Herrich-Schäffer is an exception but may
be primitive; see Macrobrochis Herrich-Schäffer); fusion of hindwing vein Sc with Rs over some distance
(usually to at least the centre of the section of Rs that forms the cell, though
the genera in the first section discussed, including Macrobrochis, show
this weakly if at all); a tymbal organ where the microtymbals are fine, shallow
grooves, each groove associated with a modified scale.
A high proportion of genera
has the valve of the male genitalia with a spine-like extension to the sacculus
that extends approximately to the apex of the main laimna.
Classification
into tribes
Bendib & Minet (1999)
attempted to segregate the subfamily into tribes, but again found the most
reliably definitive characters were in the larval morphology. For reasons given
in the previous section, practical alternatives to adult morphology are needed
to help apply this system to the diverse fauna of the tropics.
However, an adult character
was used to distinguish three of the tribes they recognised from the rest (the
character has not been investigated for the Acsalini, a monotypic tribe from
Alaska and the Yukon Territory): the condition of the apodemes of the sternite
of A2. However, the interpretation of Bendib & Minet (1999: 243) of the
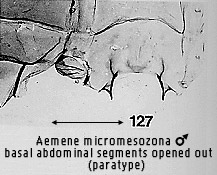
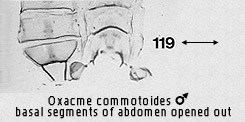
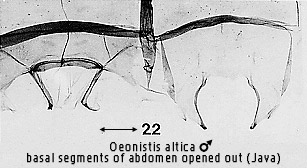
polarity of this character is unclear. In one state (Figs 119, 127, 271, 278,
298, 521 the apodemes are long, usually slender, with anterolateral processes
fused to the sternum; in the other (Figs 22, 121), the apodemes are short,
usually robust, with more articulation of the anterolateral processes with the
sternum. The second state is regarded as apomorphic by Bendib & Minet, but
stated also to occur in all non-lithosiine arctiids as well as the Lithosiini,
Endrosini and Phryganopterygini. The first state, regarded as plesiomorphic, is
restricted to the lithosiine tribes Eudesmiini, Cisthenini and Nudariini. Thus
the suggested polarity could equally well be reversed, with long, slender
apodemes being apomorphic. As will be seen, the Cisthenini and Nudariini share
the apomorphic feature of general spining in the corpus bursae, as distinct from
the pair of signa seen in most Lithosiini, the Syntominae and many Arctiinae. A
single signum, often amid more general fine scobination, is seen in the first
sequence of genera discussed below and also in the Eudesmiini (New World) and
the Phryganopterygini (Madagascar; monogeneric) where it is elongate and
scobinate (also in some of the Oriental �Agylla� group, such as Hesudra
Moore and Ghoria Moore; see Hesudra
Moore Gen. rev.). However, general spining is seen in
females of the Endrosini (Palaearctic to Oriental but not in Sundaland or
eastwards), though it also tends to be concentrated in a large, broad signum (Bendib
& Minet, 1999). The Cisthenini and Nudariini also share sclerotisation of
the ductus ejaculatorius (best developed in groups of the former), and enlarged
paraptergal sclerites (see Cisthenini again, much more developed in the
Cisthenini).
|

|
 |
|
|
|
Only three of the seven
tribes of Bendib & Minet (1999) have been clearly identified in the Bornean
fauna: the Lithosiini, the Cisthenini and the Nudariini, and appreciation of a
broader range of taxa within these may lead to their redefinition, e.g. the
Cisthenini as discussed (see Cisthenini). All will be defined and discussed in the
systematic section.
It is possible that the
first six rather plesiomorphic genera discussed may be related to the
Phryganopterygini, though several of the shared characters appear
plesiomorphic,
such as strong to moderate ocelli, little fusion of Sc and Rs in the hindwing,
R1 not anastomosing with Sc in the forewing, and no loss of radial sector or
medial veins. More unusual features shared with at least some of the Oriental
genera are the forewing areole (seen in the �Agylla� group of genera
but also true Neotropical Agylla Walker) and the longitudinal signum
mentioned above. The hindwing venation is also comparable, with all veins
usually present from Rs to CuA2.
A number of genera could not
be assigned with confidence to any of the three tribes, some because they were
represented by only limited material, but mostly because, whilst showing
distinct modifications, they did not share any of the definitive features of the
tribes. The Eugoa Walker group of genera is held together loosely by
characters of venation and female genitalia, but there is no simple feature
found unambiguously in all taxa. Of the remainder, three genera have distinct
signa, one in the case of Narosodes Moore and two in each of Tampea Snellen
and Darantasia Walker, and the others have general spining or lack it,
though females of a few (e.g. Blavia Walker) are unknown.
Biology,
ecology and biogeography
The larvae, as mentioned
above, have mostly been recorded as feeding on most groups of lower plants:
lichens, algae, mosses and liverworts. They tend to graze on these wherever they
are abundant, such as on moist walls, cliffs and rocks, the trunks and branches
of trees and, in very humid biotopes, on leaves. The last locations may
frequently have led them to be associated with the tree or plant concerned, and
the identity of this gets propagated in the literature as the host-plant, e.g.
perhaps many of the records in Yunus & Ho (1980) and Zhang (1994). But it is
apparent (Piepers & Snellen, 1904; Sevastopulo, 1940; Issiki et al., 1965;
de Worms, 1975; Rawlins, 1984; Carter & Hargreaves, 1986; Sugi, 1987;
Common, 1990; Kitching & Rawlins, 1999; Bendib & Minet, 1999) that
feeding on higher plants is rare in the group, but noted particularly in the
Lithosiini and Nudariini, where withered or living foliage may be eaten. Henwood
(1997) observed that several British species that overwinter as larvae may pass
through double the number of instars seen in species that don�t (10-12 instead
of 5-6). Numbers of instars have been observed to increase under adverse
conditions in other Lepidoptera groups (Common, 1990: 32).

Table
1. Percentage of species for Lithosiinae groups amongst various
biogeographic and ecological categories as discussed in the text.
The genus Darantasia (see Darantasia
Walker) may be a specialist feeder on Rubiaceae; if confirmed, this would be a very
unusual feature. As noted above, the genus is bisignate and so could be an early
offshoot in the subfamily. The patterning of the wings and other morphological
features are unusual. Nishihara (1992a, b) also recorded two species in the
Nudariini feeding on living higher plant leaf tissue.
Pupation is usually in a
cocoon that incorporates the larval secondary setae, and these may also be
incorporated in a palisade around the cocoon or in an open net at a distance
from where the pupa is suspended, Cyana Walker being a classic example of
the latter (see Cyana Walker). The time from formation of the cocoon to emergence of the
adult appears to be remarkably short in Sundanian species, often just a week (Piepers
& Snellen, 1904).
Most adults are nocturnal and come to light, though day-flying is noted
in the Scaptesyle Walker quartet of genera (see
Scaptesyle Walker) and sporadically in
most of the tribes of Bendib & Minet (1999).
The group shows some promise for environmental monitoring. It is diverse
and, despite the possibility of some uniformity of the larval diet across
habitats (moisture and shelter from insolation being potentially critical
factors), shows some indication of specificity to forest type, even within the
Bornean lowlands (Holloway, 1984a, 1985, 1998a). Indeed, three closely related
species in Euconosia Watson (see
Euconosia Watson) appear to segregate on lowland
forest type. Lithosiines are moderately diverse at middle altitudes but only a
few species are characteristic of elevations above 2000m. These tend to be drawn
more from the tribes Lithosiini and Nudariini than from the Cisthenini as
illustrated by the matrices of habitat preference against biogeographic range
type in Table 1. This differential representation with altitude is reflected in
a similar response to latitude; for example, the Lithosiini and Nudariini
predominate in the European fauna, joined by members of the more strictly
temperate Endrosini.
Response to disturbance is also quite marked. The survey of lowland
softwood plantations by Chey (1994; see also Chey, Holloway & Speight, 1997)
showed that many Lithosiinae can persist in these managed systems, some becoming
quite abundant. The Nudariini appear to be particularly successful, with many
species of the genera Cyana Walker, Lyclene Moore and Barsine Walker
being common. In the Lithosiini a few Brunia Moore, Macotasa Moore
and Teulisna Walker persist, and several species of Eugoa Walker
are also represented. In Seram, Holloway (1993, 1998a) found that a small number
of species, particularly a member of the nudariine genus Cyme Felder and
two Lithosiini, persisted in areas disturbed by cycles of shifting cultivation.
In open, farmed habitats representation is much lower, usually only one or two
species.
Representation in mangrove is also typically low, with Brunia antica Walker
the only Bornean species commonly encountered in this habitat; this species also
occurs on some remote islands such as the Chagos group in the Indian Ocean where
Lithosiinae are otherwise unrepresented. Eugoa mangle sp. n. and Stictane
muara sp. n. are rarer species that may be associated with mangrove.
The biogeography of the group as a whole is indicative of poor dispersal
ability. In the course of this work, apparently widespread species such as Oeonistis
altica Linnaeus have proved to be complexes of geographically more localised
species. Only Lobobasis niveimaculata Hampson and Trischalis subaurana
Walker range widely through the Indo-Australian tropics, with just two other
species, "Eilema" prabana Moore and "Eilema" monochroa Turner,
found on both sides of Weber�s Line, the major discontinuity between Oriental
and Australasian faunas in the region (Holloway, 1998b). Several genera appear
to be endemic to Sundaland (e.g. Garudinistis Hampson, Stictosia Hampson,
Euconosia Watson, Malesia van Eecke) or even Borneo (e.g. Mantala
Walker, Cyclosodes Hampson, Omiosia Hampson, Quadrasura Gen.
n., Nudariphleps Gen. n.), though all of the latter
category are rare, monobasic, and will possibly prove to occur elsewhere in
Sundaland. The monobasic Neoduma Hampson is only known from the
Philippines and Borneo, and Graptasura Hampson is found only in Sulawesi
and Borneo, one endemic species in the former and two in the latter.
|