|
The Uraniidae have recently been the subject of detailed morphological
investigation by Lees & Smith (1991) and Minet (1994[5]), extending previous
work by Sick (1937) and Minet (1983, 1986, 1991): see also reviews by Common
(1990) and Scoble (1992).
The most striking, definitive feature of the family is the occurrence of
sexual dimorphism in the position of the tympanal organs, as well as their
general structure. Whilst those of females are in a similar position to those of
the Geometridae, ventrally at the base of the abdomen, those of the male occur
laterally between the second and third tergites (Figs 159-168). Minet (1994[5])
indicated that, whilst the cuticular components of these organs in males and
females are not homologous, the sensory components (scoloparia with only two
scolopidia instead of four) may well be. The subfamily Auzeinae has a somewhat
different structure in the male tympanal organ to that of the other uraniid
subfamilies, leading Minet (1994[5]) to suggest it was the most primitive
lineage of the group (See Auzeinae).
The forewing venation provides another family characteristic, lacking
the areole seen in numerous lineages of the Drepanoidea and Geometridae, and
having R3+R4 as the terminal bifurcation of the Rs system, with R5 well separate
from this system, sharing a common stalk, or, more rarely, being connate with M1
(Holloway, 1986a: fig 23). R2 may arise independently from the cell or branch
off basally from R3+R4. However, some Epicopeiidae have a similar venation
system in the forewing (See Epicopeiidae). There is also a diagnostic sclerite in the
metathorax (Minet, 1983; Lees & Smith, 1991), though this is absent from the
Auzeinae (Minet, 1994 [5]).
The moths are generally of delicate build with deep triangular forewings
and, with the exception of the Auzeinae, tailed or angled hindwings. The veins
associated with these tails vary from subfamily to subfamily, tending to be Rs
and M3 in the Epipleminae, M3 in the Microniinae, M1 and M3+CuA1 in two-tailed
Uraniinae and M3 in single-tailed genera. Forewing pattern is usually repeated
on the hindwing except in some Epipleminae, though in all groups the hindwing is
patterned. Chaetosemata are large and ocelli are absent.
The male genitalia usually lack a gnathus. The female has a rounded
corpus bursae with a single signum, weakly or strongly spined, in a central to
subbasal position.
The eggs are domed, spherical with projecting ribs, deposited in an
upright position. The larvae usually lack secondary setae and have the primary
setae on raised pinacula. The pupal cremaster has four pairs of hooked shaftlets,
though these may be reduced in some genera (e.g. to two in Acropteris Geyer).
The relationship of the subfamilies in Fig 1 was suggested by Smith
& Lees (1991) and Minet (1994[5]). The non-auzeine subfamilies share the
metathoracic sclerite referred to above, and strong modification of the
anterolateral areas of the male third abdominal tergite.
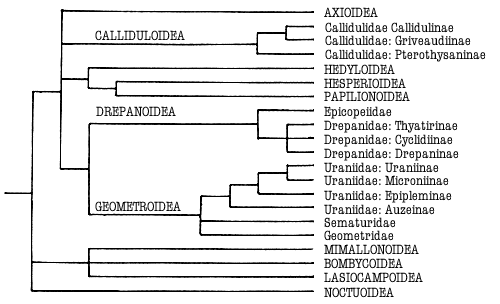
Figure 1. Phylogeny
of the Macrolepidoptera as suggested and Minet (1991)
and Kristensen & Skalski (in press). Superfamilies are indicated in
capitals. Groups treated in this work are indicated in bold.
The sister-relationship between the Uraniinae and Microniinae is
indicated by Minet (1994[5]): loss of the frenulum and retinaculum with
development of amplexiform wing coupling (a weak humeral vein may be present);
the condition of the mesothoracic presternum, with fairly short ventral arms,
the membrane between them vestigial; a short scoloparium in the male, attached
to the convex anterior area of the tympanum, the posterior part being covered
with many fine, parallel strigae. In addition, the hindwing has only one anal
vein (though this condition also occurs in the Auzeinae and some Epipleminae),
and the wing pattern usually involves strong transverse striae.
Males in some species of both the genera discussed below (the type
species of Decetia Walker, its close relatives and D. subflavata Warren;
the new species of Brachydecetia nom. n.) have CuA2 in the hindwing
arising from the cell in an unusually basal position.
There is some host-plant specialisation within the subfamilies (Lees
& Smith, 1991), with Uraniinae only reliably recorded from Euphorbiaceae and
Microniinae from Asclepiadaceae. The Epipleminae exploit a wider range of
families, but there is specialisation at a generic level as discussed later. The
only records for the Auzeinae are from the Olacaceae and the related family
Opiliaceae.
|
