|
Maurilia
iconica
Walker
Anomis iconica Walker, 1857 [1858], List Specimens lepid. Insects Colln
Br. Mus., 13: 992.
Briarda
cervina
Walker, 1866, List Specimens lepid. Insects Colln Br. Mus., 35: 1968.
Anomis candida
instabilis
Butler, 1889, Illust. typical Specimens lepid. Heterocera Colln Br. Mus., 7: 72.
Maurilia bifascia Gaede, 1915, Int. ent. Z., 9: 80.
Maurilia iconica ab.
iconicoides
Strand, 1915, Arch. Naturgesch., 80 (A2): 166.
Maurilia
tunicata
Swinhoe, 1918, Ann. Mag. nat. Hist. (9), 2: 71.
Maurilia
undaira
Swinhoe, 1918, Ann. Mag. nat. Hist. (9), 2: 71 (partim; fig. 3A, slide no. 16815).
Maurilia
fortis
Swinhoe, 1918, Ann. Mag. nat. Hist. (9), 2: 72.
Maurilia
dalama
Swinhoe, 1918, Ann. Mag. nat. Hist. (9), 2: 72.
Maurilia
gilva
Swinhoe, 1919, Ann. Mag. nat. Hist. (9), 3: 317.
Acontia
elima
Swinhoe, 1919, Ann. Mag. nat. Hist. (9), 3: 318.
Maurilia
iconicoides
Gaede, 1938, Gross-Schmett. Erde, 11: 441.
Maurilia
subiconica
Kobes, 1997: 172, syn. n.
Diagnosis and taxonomic note. Both Bornean species of this genus have very
similar and variable facies, with diffusely fasciated and variable grey to
reddish forewings, sometimes with a crescent-shaped reniform stigma. The only
potentially reliable distinction noted within Sundaland is that the posterior
half of the postmedial row of dots is slightly irregular in iconica and
straight in the next species. The species are best distinguished on male
abdominal characters (females appear to be uninformative), as indicated by Kobes
(1997) in Sumatra, recognising also a third species. The three species were also
recognised by Swinhoe (1918: loc. cit.), who described further
Australasian taxa based on females. All available Indo-Australian names
currently in the genus were placed as synonyms of iconica by Nielsen
et al. (1996), who also considered the infrasubspecific names of Strand
listed had been validated by Gaede (1938, Gross-Schmett. Erde, 11: 441),
though Gaede�s account is confusing, stating that all forms were from India,
implying variability rather than segregated races (though the type material of
rufescentibrunnea is from Singapore and that of iconicoides from
New Guinea).
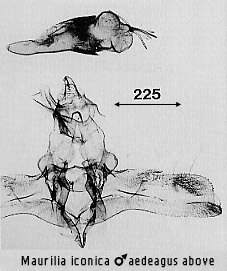
In
iconica the basal hair pencils of the abdomen are larger than in the
other two species and blackish rather than pale brown. In the genitalia the
central costal process of the valve is short, blunt. The aedeagus vesica has a
cluster of four to six blade-like cornuti at the apex, directed dorsally or
radiating. Lateral setal patches on the juxta/anellus have the setae moderate in
thickness, some relatively long. The Swinhoe taxa fortis, dalama,
and gilva are based on females that are probably referable to iconica.
Material from Seram eastwards is generally a darker, richer red than in typical
Oriental iconica, and could be referred to ssp. tunicata Swinhoe.
The third species,
M.
cervina
Walker stat. rev. (=
instabilis
Butler, syn. rev. and
pallidipennis
Warren, syn. n.) is paler and greyer than either of the Bornean species.
The costal process of the valve is smaller than in iconica, almost
vestigial. The aedeagus vesica has more slender, ventrally directed, curved
cornuti apically. The lateral setal patches of the juxta/anellus are more
dispersed and robust than in iconica. The species occurs in India and
Burma and may be that from Sumatra referred to the African M. arcuata
Walker by Kobes (1997); arcuata lacks cornuti in the vesica whereas the
Sumatran species is stated to have three slender ones.
The Australian taxon elima Swinhoe, placed as a synonym of iconica
in Nielsen et al. (1996), is based on a single female that appears
identical to M. arcuata ab. lubinata Strand. Its status needs
further investigation.
Geographical range. Indo-Australian tropics from Sri Lanka to Queensland,
Samoa, Rarotonga and New Caledonia.
Habitat preference. Only one Bornean male has been definitely identified by
dissection: from lowland alluvial forest during the Mulu survey. Two possible
females were taken in lowland forest in Brunei. All were taken below 100m.
Biology. Bell (MS) described larvae in India attributed to iconica;
Gardner (1941, 1946b) described the chaetotaxy. The larva has segments T1-T3
swollen into a berry-like tumidity. This is much more conspicuous in early
instars. The body narrows to A8 which bears dorsally a pair of rounded, conical
tubercles. All four prolegs are present, equal. The colour is a dark brown,
profusely dotted and marbled with lighter brown. There are broken, thin, white
subdorsal, dorsolateral, lateral and spiracular lines, though these are
indistinct on the berry, which is more a dull olive green with white surrounds
to the bases of the black setae. Between the lines the body may be tinged more
greenish or orange, though there is some variation in colouring and marking,
with shades of green rather than brown, and yellow rather than white. This
variability may indicate more than one member of the complex was studied.
The pupa is enclosed in a cocoon of whitish silk, a dirty fuscous, boat-shaped, semiovoid in the midst of silken webbing on the face of a curled leaf or two
adjacent ones. The pupa is claviform, with the abdominal end narrow but bluntly
rounded without a cremaster. Segments A9 and A10 are minutely rugose dorsally in
a sort of honeycomb where the longitudinal ridges are much stronger. Sound
production is not noted.
The larvae live on the undersides of young leaves or leaf stalks when older, and
retire to twigs or branches, dropping off on a silk when disturbed.
Recorded host-plants were Vatica (Dipterocarpaceae) and Terminalia
(Combretaceae). Mathur (1942) noted the larva also on Shorea (Dipterocarpaceae;
also in unpublished FRIM records), Anogeissus (Combretaceae) and
Tectona (Verbenaceae). Yunus & Ho (1980) gave Saccharum (Gramineae)
as a host-plant. A definite host association for iconica from Samoa
(Robinson, 1975) is Terminalia (Combretaceae).
<<Back
>>Forward <<Return
to Content Page
|