|
The Aganainae are a group that has often been treated as a full family
(e.g. Inoue et al., (1982), as Hypsidae) or as a subfamily of the
Arctiidae (e.g. Watson, Fletcher & Nye, 1980). Watson, Fletcher & Nye
(1980) and Kitching (1984) have discussed the nomenclature and taxonomic history
of the group.
The condition of the counter-tympanal hood has been mentioned earlier;
the spiracle occurs in an ambiguous position between two weak structures that
may represent the counter-tympanal hood. There is no tymbal organ; absence
cannot be convincingly presented as due to loss (Kitching, 1984). The larvae of
typical Aganainae have a single subventral seta on the mesothoracic and
metathoracic segments, a noctuid feature, but lack the ventral prothoracic gland
of the noctuids. Kitching (1984) suggested, on these criteria, that the group
might require reinstatement as a full family. Minet (1986) referred to
characters of the basal abdominal sternite and tympanal organs that were noctuid-like.
There are additional features that might indicate that the group should
be placed with the Noctuidae.
The forewing pattern includes an orbicular stigma in the forewing cell.
This is a frequent feature in Noctuidae but is not seen in other noctuoids
except possibly some Notodontidae (e.g. Phalera). It can be seen most
clearly on the underside of the wing but is often obscured on the upperside.
The labial palps are ascending, with a long, slender terminal segment
(Fig. 147) as seen in numerous genera of catocaline noctuids (e.g. Othreis).
Features of the male abdomen, such as a single corema on the eighth
abdominal sternite and, to some extent, the complexity of the lobing of the
aedeagus vesica, are also noctuid-like.
The aganaine genera represented in Borneo mostly fall into a probable
monophyletic group, the exception being Agape Felder. The component taxa
show varying development of coremata on the male eighth segment; a basal central
one on the sternite and a lateral terminal pair; in Agape there is a
strong central one only. The valves in the main group have a strong terminal
spine or harpe to the sacculus; in Agape the sacculus is not clearly
differentiated from the rest of the valve and there is no harpe. In the female
genitalia there is a pair of finely scobinate secondary lobes ventrally between
the ovipositor lobes.
All genera have dark banding or maculation on the abdomen. All have a
subbasal black spot on the tegulae, towards the rear of the thorax and some
pattern elements with small black spots at the base of the forewings. In genera
apart from Agape there are usually taxa where this black spotting
delimits a zone of orange colour. The retinaculum of the male is slender, long,
apically hooked in the main group of genera, but absent in Agape where
the frenulum lodges in the tuft of scales arising from CuA that usually retains
the frenulum in the female.
The male antennae are ciliate, filiform or slightly bipectinate (some Asota),
the pectinations being very slender and strongly curved, invested with a
basal fan of fine cilia.
All of the main group of genera have a slight elliptical pucker to the
wing membrane on the dorsum subtornally; on the underside this bears a small,
dense ridge of pale orange scales on its anterior margin. In extreme instances
the dorsal margin is slightly convex at that point.
A cladogram of relationships is presented in Fig. 7. Apomorphies
defining each grouping are discussed in the generic descriptions following. The
main group, excluding Agape, is defined by: the forewing dorsal pucker;
black spots on the tegulae and base of the forewing; the array of coremata on
the male eighth sternite.
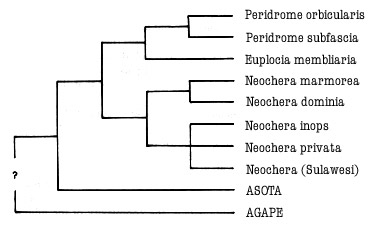
Figure
7. Diagram
of relationships within the Aganainae.
Host-plant records for Asota are predominantly from Ficus (Moraceae).
The two records located for the sister-group are from the Apocynaceae and
Asclepiadaceae.
The mature larvae (Gardner, 1941) show no reduction of prolegs, unlike
many �semi-looper� catocalines. The crochets of the prolegs are in a
homoideous, uniordinal mesoseries. The larvae appear sparsely hairy because of
the great length of the primary setae; these arise from chalazae. There are no
secondary setae.
>>Forward
<<Return to Contents page
|
