|
The epiplemines are very much smaller than most members of the other
subfamilies. The majority have hindwings with tails at Rs and M3, the fasciation
is often sharply angled, the angle more or less in line with the more posterior
tail. The submarginal zone of the forewing is often darker distal to an
irregularly lunulate or arcuate boundary. Some genera have vein M2 in the
hindwing weak or absent. The costal margin of the hindwing is often strongly
excavate centrally. There are usually two anal veins compared with one in other
subfamilies (Common, 1990) (but see Chionoplema Gen.n
and Monobolodes
Warren for exceptions).
The male antennae are frequently lamellate, often also uniserrate. In
some genera they are strongly bipectinate. The strip separating the male
tympanum from the counter-tympanum is diagnostically sclerotised, rather than
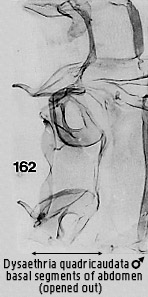
membranous (Minet, 1983, 1994[5]). Male tympana are illustrated in Figs
162 and 163, and female ones in Fig 165. A frenulum is present (Scoble, 1992).
There are no really definitive features of the male genitalia, though
the base of the valve usually supports one or two hair pencils. The transtillae
sometimes form a bridge between the valves. Several genera prove exceptions to
the observation of Common (1990) that uraniids lack a gnathus. In addition to
the Bornean genera where this is stated to occur, a gnathus is found
in the Neotropical genus Psamathia Walker, the Australian genus Dysrhombia
Warren, and the Oriental (continental) genus Eversmannia Staudinger.
It is frequently divided though in Eversmannia it is entire, distally
trifid.
The larval head is hypognathous. The body cuticle is finely spinulose,
and the primary setae arise from prominent pinacula or chalazae: there are no
secondary setae (Common, 1990). Common noted the crochets in the larvae of Phazaca
to be uniordinal in anterior and posterior curved bands on each proleg. In
general, the crochets form a strongly curved mesoseries, resembling a penellipse
and the prolegs have strongly sclerotised lateral plates (Holloway, Bradley
& Carter, 1987).
The pupal cremaster has four pairs of hooked shaftlets, the same
groundplan state as in the other uraniid subfamilies and geometroid groups, but
often the most distal pair is enlarged, with reduction or loss of the others.
Pupation is usually in a cocoon.
Lees & Smith (1991) noted that host-plants recorded for the
subfamily overlapped almost completely the much wider range recorded for the
Sphingidae (see also Holloway (1987)). However, there appears to be a much
stronger trend for specialisation within epiplemine genera or species groups
within genera. Only Dysaethria quadricaudata Walker (Dysaethria
quadricaudata Walker comb. n.
- Dysaethria scopocera
Hampson comb.n. longiductus ssp.n) is
definitely polyphagous though within the Rubiaceae. Plant families exploited by
Bornean genera are Annonaceae, Bignoniaceae, Caprifoliaceae, Daphniphyllaceae,
Oleaceae, Rubiaceae, Scrophulariaceae and Verbenaceae. Only the Rubiaceae and
Daphniphyllaceae are recorded for more than one genus. In addition to the
species discussed here, "Epiplema" latifasciata Moore was noted as
feeding on Fagraea ("Fagiaea", Loganiaceae) in the original
description.
The resting posture of the adults is sometimes modified, particularly in
Phazaca Walker, Monobolodes Warren and two new genera (below).
The wings are otherwise held flat against, or parallel to, the substrate at
rest.
The subfamily is pantropical, extending only weakly into temperate zones
(e.g. in China and Japan). However, it has a much greater representation in
montane zones than do the other subfamilies. The group appears to be
proportionally more diverse in Australasian tropical faunas, particularly that
of New Guinea, than in Oriental ones (Holloway, 1993), though it does not extend
further east than Samoa. Janse (1932) described the southern African fauna and
Boudinot (1982) that of Madagascar.
The generic classification requires extensive revision, and an attempt
has been made here to rationalise the situation in the Indo-Australian tropics.
Application of the genus-group name Epiplema Herrich-Schäffer, based on
a robust Neotropical species, E. acutangularia Herrich-Schäffer, to
Indo-Australian taxa is inappropriate, though a few species listed at the end of
the account below have yet to be found better placements.
>>Forward
<<Return to Contents page
|